استفاده از مدلهاي جانوري براي مطالعه بيماريهاي انسان
موشهای ترانسژنی هدفگذاری شده با استفاده از سلولهای بنيادی رویانی(ES)
روش جديدتر و معتبرتري که براي توليد موشهاي ترانسژن به کار گرفته شده عبارت است از وارد کردن يک نسخه ژن بهيک جايگاه هدفگذاري شده از ژنوم موش توسط نوترکيبي همولوگ. ويژه بودن اين فرايند، توليد موشهاي ترانسژنيک را که از جهشهاي متناظر در ژنهاي کلون شده بيماري انسان به وجود آمدهاند، ميسر ميسازد. اين روش با ايجاد شيوهاي براي کشت سلولهاي ES چند قوهزا در خارج از بدن و حفظ چند قوهزايي آنها حتي بعد از چندين تقسيم سلولي امکان پذير شد.
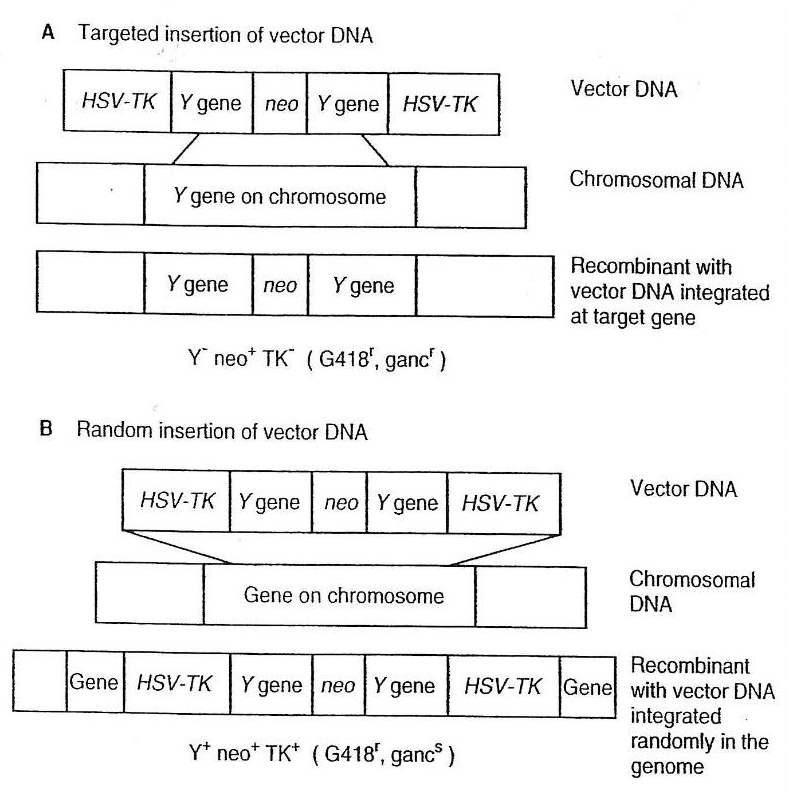
در اين روند، سلولهاي ES موش از توده دروني بلاستوسيت نژاد ۱۲۹/SvJ کونژنيکموشهاي قهوهاي برداشته شدهاند. سلولهاي ES در محيط کشت رشد داده شدهاند و سپس با حاملي که جهت راه اندازي نوترکيبي با توالي ژن همولوگ در ژنوم موش ساخته شده بود، ترنسفکت شدهاند. حامل، تواليهاي DNA ژن را که بايد هدفگذاري شوند به همراه يک ژن مارکر مثبت و يک ژن مارکر منفي حمل ميکند. مارکرها ميتوانند در روند گزينش مثبت-منفي به کار روند و جداسازي سلولهاي ES که حامل DNA را به يک جايگاه ويژه وارد کردهاند ميسر سازند.
هر سلولي که با DNA حامل ترکيب ميشود، چه از طريق ادغام تصادفييا توسط نوترکيبي همولوگ در جايگاه هدفگذاري شده، ژن neo(مارکر مثبت) را در خود ادغام کرده و نسبت به آنتي بيوتيک G418 مقاوم ميشود.
سلولهاي ES گرفته شده از يک موش قهوهاي، در شرايط آزمايشگاه کشت داده شده و با استفاده از روند هدفگذاري که يک ژن ويژه را غيرفعال ميسازد، به طريق ژنتيکي اصلاح شدهاند. سلولهاي اصلاح شده ES به درون بلاستوسيست يک موش سياه تزريق ميشود و بلاستوسيست به يک مادر اجارهاي پيوند ميشود و تا پايان رشد ميکند. موشهاي کايمريک قهوهاي-سياه، موشهايي هستند که از هر دو نوع سلول به وجود آمدهاند. آزمايشهاي جفتگيري، موشهاي کايمريک نر که داراي ژن تغيير داده شده به طريق ژنتيکي بودند را در لایهزاینده، شناسايي ميکنند زيرا آنها در آميزش با مادههاي سياه، موشهاي قهوهاي به وجود ميآورند. جفتگیری دو موش هتروزيگوت، يک موش ترانسژني يا ناکاوت را به وجود ميآورد که در ژن معيوب هموزيگوت است.
سپس سلولهاي مقاوم در برابر G418 دريک محيط کشت حاوي گانسيکلووير قرار داده ميشوند. گانسيکلووير سلولهايي را که به صورت اتفاقي با ژن تيميدينکيناز HSV(مارکر منفي) ادغام شدهاند را ميکشد. ادغام تصادفي، که در شکل B9-14 نيز نشان داده شده است، صورت متفاوتي از نوترکيبي را به غير از نوترکيبي همولوگ شامل ميشود. وقتي DNA حامل ادغام تصادفي را انجام ميدهد، از پايانههايش وارد DNA ژنومي ميشود. مارکر منفي يا ژن تيميدينکيناز HSV، در پايانههاي DNA حامل قرار دارد. در طول فرايند ادغام تصادفي، اين ژن ادغام شده و به بخش ثابتي از DNA ژنومي تبديل ميشود. در مقابل، در طول فرايند نوترکيبي همولوگ، پايانههاي DNA حامل که حاوي ژن تيميدينکيناز HSV هستند، وارد ژنوم نشده و از دست ميروند. از اين رو، سلولي که در جايگاه هدف نوترکيبي همولوگ را انجام ميدهد، فاقد ژن تيميدينکيناز HSV است و در برابر گانسيکلووير مقاوم ميباشد. در حاليکه سلولي که دخول تصادفي را انجام ميدهد داراي ژن تيميدينکيناز HSV است و به عمل کشتن گانسيکلووير حساسيت نشان ميدهد. استفاده از روش گزينش مثبت-منفي، جداسازي سلولي که يک توالي ژني ويژه در جايگاه هدفش ادغام کرده را از هزاران سلول ديگر ميسر ميسازد.
وقتي سلول ES مناسب از طريق گزينش مثبت و منفي جداسازي شد و مشخص گشت کهيک ژن ويژه غيرفعال شده دارد، در محيط کشت تکثير ميشود تايک جمعيت از سلولهاي ES دگرگون شده به طريق ژنتيکي توليد شود(شکل ۸-۱۴). سلولهاي ES تغيير داده شده به طريق ژنتيکي به بلاستوسيت به دست آمده از يک دسته موش با رنگهاي مختلف، ريزتزريقشدند(مثلا موش C57/6J یا موش سياه). ژن پوشش سياه نسبت به ژن پوشش قهوهاي مغلوب است و موشهاي کايمريک را به وجود ميآورد. سپس بلاستوسيت به رحم يک مادر اجارهايسياه پيوند ميشود و اجازه داده ميشود که فرزندان حاصله تا پايان رشد کنند.
هر موش تازه متولد شدهاي که داراي پوشش کايمريک يا ترکيبي سياه و قهوهاي باشد، نشان ميدهد که سلولهاي ES زنده ماندهاند و در اين موش تکثير شدهاند. موشهاي سياه نشان ميدهند که سلولهاي ES در آنها وجود ندارند. با اين حال، شناسايي موشهاي کايمريکي که جهش هدفدار در سلولهاي زايايشان رخ داده ضروري ميباشد. براي اين شناسايي بايد موشهاي کايمريک نر را با موشهاي سياه ماده جفت کرد و منتظر موشهاي قهوهاي شد که با اسپرمي که از سلولهاي ES اوليه تغييريافته به طريق ژنتيکي مشتق شده، به وجود ميآيند.
موشهاي قهوهاي با تکنيکهاي مولکولي بررسي ميشوند تا مشخص شود که آيا آنها يک نسخه از ژن جهشيافته را در جايگاه ژنومي مناسب دارا هستند يا خير. در مورد ژنهايي که روي کروموزوم X قرار نگرفتهاند، فقط يک آلل ژن توسط نوترکيبي رخ داده بين DNA حامل و کروموزوم ميزبان، غيرفعال ميشود. موش هايي که داراي دومين آلل غيرفعال نيز هستند در جفتگيري بعدي انتخاب ميشوند. از جفتگيري بين موشهايي که در ژن جهشيافته هتروزيگوتاند، فرزنداني حاصل ميشوند که با فراواني ۲۵% در ژن جهشيافته هموزيگوت هستند. اين موشهاي هموزيگوت ناکاوتبه عنوان سيستمهاي مدل به کار گرفته ميشوند تا از طريق آنها عواقب فنوتيپي جهش در يک ژن ويژه بررسي شود. اين تکنيک که ممکن است تکميل آن تقريبا يک سال طول بکشد، براي توليد صدها موش ترانسژن که فاقد ژنهاي مختلف بيماريهاي انسان هستند، به کار ميرود.
تو ضیحات مربوط به شکل: هدف گذاری ژنها با استفاده از سیستم گزینش مثبت-منفی A)يک حامل هدفگير دربردارنده مارکر انتخابي مثبت(ژن neo) که ميان تواليهاي DNA ژني که بايد هدفگیری شوند(ژن Y) و مارکر منفي که ژني است که در ويروس هرپس سيمپلکس(HSV) است که تيميدينکيناز را کد ميکند و در دو انتها قرار دارد، قرار گرفته است. در مدت ورود حامل به درون ژن Y هدفگذاري شده توسط نوترکيبي همولوگ، دو انتهاي DNA حامل از دست ميروند و سلول حاصل در ژن Y معيوب است)ֿY) و داراي ژن neo(neo+، مقاوم در برابر(G418)) و فاقد ژن TK(ֿTK، مقاوم در برابر گانسيکلووير) ميباشد.B)در طول فرايند دخول تصادفي DNA حامل به کروموزومهاي ميزبان از طريق نوترکيبي غيرهمسان، DNA حامل از دو انتهايش وارد ميشود. سلول به وجود آمده، در Y فعال(+Y) است و داراي ژن neo (neo+، مقاوم در برابر G418) و داراي ژن TK(TK+، حساس به گانسيکلووير) ميباشد.